

Reprodução proibida. Art.184 do Código Penal e Lei 9.610 de 19 de fevereiro de 1998.
17 7
Capítulo 6 • Citoplasma e organelas citoplasmáticas
2 Complexo golgiense
Muitas proteínas produzidas pelos ribossomos do retículo granuloso são enviadas diretamente
para outra estrutura membranosa citoplasmática: o complexo golgiense, ou complexo de Golgi
(ou ainda aparelho de Golgi). Esse componente citoplasmático é constituído por de 6 a 20 bolsas
membranosas achatadas, denominadas cisternas ou vesículas, empilhadas umas sobre as outras.
Nas células vegetais, há conjuntos de cisternas dispersos pelo citoplasma, sendo cada conjunto
denominado golgiossomo ou dictiossomo. Nas células animais, as cisternas do complexo gol-
giense se concentram em uma região específica do citoplasma, geralmente próxima do núcleo e
de um par de estruturas tubulares conhecidas como centríolos, que estudaremos mais adiante.
Nas cisternas do complexo golgiense, certas proteínas produzidas no retículo granuloso são qui-
micamente modificadas pela adição de glicídios, processo denominado glicosilação de proteínas.
É também no complexo golgiense que ocorre a síntese de determinados carboidratos.
Secreção celular
A maioria das proteínas que atuam no ambiente externo à célula, como as enzimas que fa-
zem a digestão dos alimentos em nosso tubo digestório, passa pelo complexo golgiense, onde
são “empacotadas” no interior de bolsas membranosas, denominadas grãos de zimogênio, para
serem enviadas aos locais extracelulares em que atuarão. O complexo golgiense é, portanto, o
responsável pela secreção celular, nome atribuído ao processo de eliminação de substâncias
úteis por células secretoras.
A microscopia eletrônica e outras técnicas citológicas revelaram como se dá a transferência
das proteínas produzidas no retículo endoplasmático granuloso para as cisternas do complexo
golgiense: ela ocorre por meio das chamadas vesículas de transição, bolsas membranosas re-
pletas de proteínas recém-sintetizadas, que surgem por brotamento na superfície das vesículas
do retículo endoplasmático granuloso. Uma vez liberadas do retículo, as vesículas de transição
deslocam-se em direção ao complexo golgiense e se fundem à sua cisterna mais externa, onde
despejam o conteúdo proteico que transportam. Essa primeira cisterna golgiense a receber pro-
teínas do retículo forma em suas bordas novas vesículas de transição repletas de proteína, as
quais se fundem à cisterna imediatamente acima. A segunda cisterna golgiense origina, por sua
vez, novas vesículas de transição que irão se fundir à cisterna seguinte e assim sucessivamente.
As vesículas liberadas pela última cisterna da pilha dirigem-se a seu destino final, que pode ser o
próprio citoplasma, no caso de a vesícula conter proteínas estruturais da membrana ou enzimas li-
sossômicas, ou o meio extracelular, no caso de a vesícula conter proteínas a serem secretadas.
Os citologistas costumam dizer que o
conjunto de cisternas do complexo golgiense
é polarizado, pois apresenta uma face na qual
as vesículas provenientes do retículo endo-
plasmático granuloso se fundem, e uma face
oposta, da qual se desprendem as vesículas em
direção a seus destinos. A face do complexo
golgiense voltada para o retículo granuloso é
chamada face cis ou face formativa. A face
oposta, em que brotam as vesículas contendo
as proteínas modificadas e selecionadas de
acordo com seu destino, é denominada face
trans ou face de maturação. (Fig. 6.7)
Figura 6.7 Representação esquemática da estrutura
do complexo golgiense parcialmente cortado para
mostrar sua organização. Note a face cis, por onde
proteínas provenientes do retículo endoplasmático
penetram no complexo golgiense, e a face trans, por
onde as proteínas modificadas e empacotadas deixam
o complexo. A ilustração mostra um instante congelado
do processo; vesículas são continuamente liberadas
pelo retículo e se fundem à face cis do complexo; as
cisternas, por sua vez, liberam continuamente mais
vesículas de transição. (Representação sem escala,
cores-fantasia.) (Baseado em Lodish, H. e cols., 2004.)
Vesículas
de secreção
Membrana
plasmática
Lisossomo
primário
COMPLEXO
GOLGIENSE
Bolsas do
complexo
golgiense
RETÍCULO
ENDOPLASMÁTICO
GRANULOSO
Transporte de
proteínas do RE para
o complexo golgiense
Vesículas
Face trans
Face
cis
Bolsas
intermediárias
Ribossomos, onde ocorre
a síntese de proteínas
Secreção


Crie sua conta grátis para liberar esse material. 🤩
Já tem uma conta?
Ao continuar, você aceita os Termos de Uso e Política de Privacidade
Duodeno
Eliminação da secreção
(enzimas digestivas)
CÉLULA ACINOSA
Complexo golgiense
(concentração e
empacotamento
das enzimas)
Retículo
endoplasmático
granuloso (síntese
das enzimas)
Ácino
Canal
pancreático
Ve sícula de
secreção
(grão de
zimogênio)
Núcleo
PÂ NCREAS
FÍGADO
Reprodução proibida. Art.184 do Código Penal e Lei 9.610 de 19 de fevereiro de 1998.
17 8
Unidade B • Organização e processos celulares
Acrossomo
Cauda
ESPERMATOZOIDE
Citoplasma
eliminado
Nas células vegetais, o complexo golgiense também desempenha função secretora. É por
meio dele que são secretadas as glicoproteínas e alguns polissacarídios (pectina e hemicelulose)
que integram a parede celular e constituem o “cimento” que une células vizinhas. Além disso,
o complexo golgiense pode originar e “abastecer” de substâncias o vacúolo central típico das
células vegetais. Vesículas liberadas do complexo golgiense se fundem continuamente a esse
vacúolo, nele lançando enzimas que atuam na digestão intracelular aí realizada.
O complexo golgiense também desempenha papel importante na formação dos espermato-
zoides dos animais, originando o acrossomo (do grego acros, alto, topo, e somatos, corpo),
uma grande vesícula repleta de enzimas digestivas, que ocupa a ponta da cabeça do
espermatozoide. As enzimas digestivas contidas na vesícula acrossômica têm por
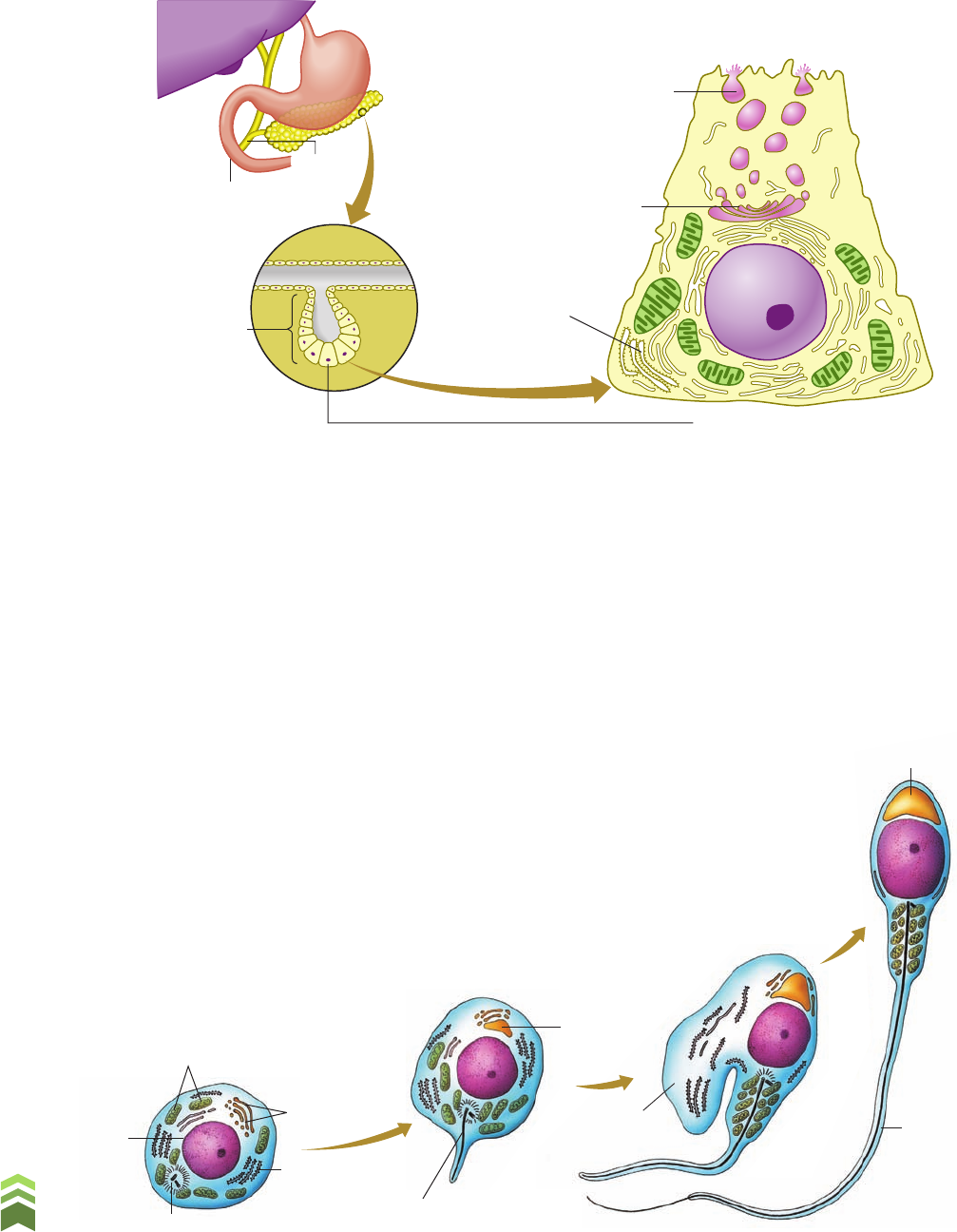
função perfurar as membranas do óvulo na fecundação. (Fig. 6.9)
Dentre tantas funções, o complexo golgiense também é responsável pela produ-
ção da organela citoplasmática denominada lisossomo, como veremos a seguir.
Figura 6.9 Representação esquemática da diferenciação do
espermatozoide. À medida que a espermátide se transforma
em espermatozoide, as cisternas do complexo golgiense
acumulam enzimas; estas se fundem e originam a vesícula
acrossômica, localizada na extremidade da cabeça do
es pe rma tozo id e. (Representação sem escala, cores-fantasia.)
ESPERMÁTIDE
Mitocôndrias
Núcleo
Complexo
golgiense
Centríolo
Retículo
granuloso
Vesícula
acrossômica
Crescimento do
centríolo
Figura 6.8 Representação esquemática da localização e da estrutura de uma célula acinosa. O pâncreas
contém inúmeras bolsas, denominadas ácinos pancreáticos, cujas paredes são constituídas por células
secretoras de enzimas digestivas (células acinosas). (Representação sem escala, cores-fantasia.)
Diversas substâncias de natureza proteica, além das enzimas digestivas já mencionadas,
passam pelo complexo golgiense para ser secretadas; é o caso de certos hormônios e de subs-
tâncias mucosas, como as produzidas por nossas vias respiratórias. (Fig. 6.8)
Crie sua conta grátis para liberar esse material. 🤩
Já tem uma conta?
Ao continuar, você aceita os Termos de Uso e Política de Privacidade

Reprodução proibida. Art.184 do Código Penal e Lei 9.610 de 19 de fevereiro de 1998.
17 9
Capítulo 6 • Citoplasma e organelas citoplasmáticas
3 Lisossomos
Lisossomos (do grego lise, quebra) são bolsas membranosas que contêm dezenas de tipos
de enzimas digestivas, capazes de digerir grande variedade de substâncias orgânicas. Dentre as
enzimas lisossômicas, destacam-se as nucleases, que digerem ácidos nucleicos (DNA e RNA),
as proteases, que digerem proteínas, as fosfatases, que atuam removendo fosfatos de nucleo-
tídios, de fosfolipídios e de outros compostos, além de enzimas que digerem polissacarídios e
lipídios. Células animais podem conter centenas de lisossomos.
A membrana lipoproteica dos lisossomos é capaz de bombear ativamente íons H1 e C,2 do
citosol para o interior da organela. Esses íons formam ácido clorídrico (HC,), o que faz o interior
do lisossomo ter um pH ácido, em torno de 4,8, ideal para a ação das enzimas lisossômicas. Essas
enzimas praticamente não atuam em pH neutro, como o do citosol e o do líquido que banha as
células. Assim, se eventualmente um lisossomo se rompe e libera suas enzimas no citoplasma,
não chega a haver autodigestão da célula, pois o pH do citosol gira em torno de 7,0 e 7,3. Um
mistério ainda não desvendado pelos cientistas é como a membrana do lisossomo consegue se
proteger de ser digerida por suas próprias enzimas.
Os lisossomos recém-produzidos pelo complexo golgiense vagam pelo citoplasma até se fundir
a bolsas membranosas contendo materiais a serem digeridos. Enquanto essa fusão não ocorre,
eles são denominados lisossomos primários, pois ainda não iniciaram sua atividade de digestão.
Quando se fundem a bolsas membranosas com os materiais que serão digeridos e suas enzimas
entram em ação, eles passam a ser chamados de lisossomos secundários.
Lisossomos podem digerir material capturado do exterior por fagocitose ou por pinocitose,
o que constitui sua função heterofágica, ou digerir partes desgastadas da própria célula, o que
se denomina função autofágica. (Fig. 6.10)
Figura 6.10 Acima, representação esquemática das funções
heterofágica e autofágica dos lisossomos. (Representações
sem escala, cores-fantasia.) (Baseado em Campbell, N. e cols.,
1999.) À esquerda, micrografia de um corte de célula animal
ao microscópio eletrônico de transmissão em que se veem
lisossomos secundários (Li), alguns deles digerindo partes da
própria célula (aumento 8.0003).
Mi
Mi
Li
Li
Li
REG
1um
Lisossomo
secundário
Lisossomo
primário
Material
sendo
fagocitado
Fagossomo
Vacúolo
autofágico
Mitocôndria
inativa englobada
Bolsas
membranosas
Lisossomos
primários
Pinossomo
Material englobado
por pinocitose
Crie sua conta grátis para liberar esse material. 🤩
Já tem uma conta?
Ao continuar, você aceita os Termos de Uso e Política de Privacidade
Prévia do material em texto
R ep ro d uç ão p ro ib id a. A rt .1 84 d o C ód ig o P en al e L ei 9 .6 10 d e 19 d e fe ve re iro d e 19 98 . 177 C a p ít u lo 6 • C it o p la sm a e o rg a n e la s ci to p la sm át ic a s 2 Complexo golgiense Muitas proteínas produzidas pelos ribossomos do retículo granuloso são enviadas diretamente para outra estrutura membranosa citoplasmática: o complexo golgiense, ou complexo de Golgi (ou ainda aparelho de Golgi). Esse componente citoplasmático é constituído por de 6 a 20 bolsas membranosas achatadas, denominadas cisternas ou vesículas, empilhadas umas sobre as outras. Nas células vegetais, há conjuntos de cisternas dispersos pelo citoplasma, sendo cada conjunto denominado golgiossomo ou dictiossomo. Nas células animais, as cisternas do complexo gol- giense se concentram em uma região específica do citoplasma, geralmente próxima do núcleo e de um par de estruturas tubulares conhecidas como centríolos, que estudaremos mais adiante. Nas cisternas do complexo golgiense, certas proteínas produzidas no retículo granuloso são qui- micamente modificadas pela adição de glicídios, processo denominado glicosilação de proteínas. É também no complexo golgiense que ocorre a síntese de determinados carboidratos. Secreção celular A maioria das proteínas que atuam no ambiente externo à célula, como as enzimas que fa- zem a digestão dos alimentos em nosso tubo digestório, passa pelo complexo golgiense, onde são “empacotadas” no interior de bolsas membranosas, denominadas grãos de zimogênio, para serem enviadas aos locais extracelulares em que atuarão. O complexo golgiense é, portanto, o responsável pela secreção celular, nome atribuído ao processo de eliminação de substâncias úteis por células secretoras. A microscopia eletrônica e outras técnicas citológicas revelaram como se dá a transferência das proteínas produzidas no retículo endoplasmático granuloso para as cisternas do complexo golgiense: ela ocorre por meio das chamadas vesículas de transição, bolsas membranosas re- pletas de proteínas recém-sintetizadas, que surgem por brotamento na superfície das vesículas do retículo endoplasmático granuloso. Uma vez liberadas do retículo, as vesículas de transição deslocam-se em direção ao complexo golgiense e se fundem à sua cisterna mais externa, onde despejam o conteúdo proteico que transportam. Essa primeira cisterna golgiense a receber pro- teínas do retículo forma em suas bordas novas vesículas de transição repletas de proteína, as quais se fundem à cisterna imediatamente acima. A segunda cisterna golgiense origina, por sua vez, novas vesículas de transição que irão se fundir à cisterna seguinte e assim sucessivamente. As vesículas liberadas pela última cisterna da pilha dirigem-se a seu destino final, que pode ser o próprio citoplasma, no caso de a vesícula conter proteínas estruturais da membrana ou enzimas li- sossômicas, ou o meio extracelular, no caso de a vesícula conter proteínas a serem secretadas. Os citologistas costumam dizer que o conjunto de cisternas do complexo golgiense é polarizado, pois apresenta uma face na qual as vesículas provenientes do retículo endo- plasmático granuloso se fundem, e uma face oposta, da qual se desprendem as vesículas em direção a seus destinos. A face do complexo golgiense voltada para o retículo granuloso é chamada face cis ou face formativa. A face oposta, em que brotam as vesículas contendo as proteínas modificadas e selecionadas de acordo com seu destino, é denominada face trans ou face de maturação. (Fig. 6.7) Figura 6.7 Representação esquemática da estrutura do complexo golgiense parcialmente cortado para mostrar sua organização. Note a face cis, por onde proteínas provenientes do retículo endoplasmático penetram no complexo golgiense, e a face trans, por onde as proteínas modificadas e empacotadas deixam o complexo. A ilustração mostra um instante congelado do processo; vesículas são continuamente liberadas pelo retículo e se fundem à face cis do complexo; as cisternas, por sua vez, liberam continuamente mais vesículas de transição. (Representação sem escala, cores-fantasia.) (Baseado em Lodish, H. e cols., 2004.) Vesículas de secreção Membrana plasmática lisossomo primário CoMPlexo golgIeNSe bolsas do complexo golgiense ReTíCulo eNDoPlaSMáTICo gRaNuloSo Transporte de proteínas do Re para o complexo golgiense Vesículas Face trans Face cis bolsas intermediárias Ribossomos, onde ocorre a síntese de proteínas Secreção Duodeno Eliminação da secreção (enzimas digestivas) CÉLULA ACINOSA Complexo golgiense (concentração e empacotamento das enzimas) Retículo endoplasmático granuloso (síntese das enzimas) Ácino Canal pancreático Vesícula de secreção (grão de zimogênio) Núcleo PÂNCREAS FÍGADO R ep ro d uç ão p ro ib id a. A rt .1 84 d o C ód ig o P en al e L ei 9 .6 10 d e 19 d e fe ve re iro d e 19 98 . 178 U n id a d e B • O rg a n iz a çã o e p ro ce ss o s ce lu la re s Acrossomo Cauda EspErmAtozoidE Citoplasma eliminado Nas células vegetais, o complexo golgiense também desempenha função secretora. É por meio dele que são secretadas as glicoproteínas e alguns polissacarídios (pectina e hemicelulose) que integram a parede celular e constituem o “cimento” que une células vizinhas. Além disso, o complexo golgiense pode originar e “abastecer” de substâncias o vacúolo central típico das células vegetais. Vesículas liberadas do complexo golgiense se fundem continuamente a esse vacúolo, nele lançando enzimas que atuam na digestão intracelular aí realizada. O complexo golgiense também desempenha papel importante na formação dos espermato- zoides dos animais, originando o acrossomo (do grego acros, alto, topo, e somatos, corpo), uma grande vesícula repleta de enzimas digestivas, que ocupa a ponta da cabeça do espermatozoide. As enzimas digestivas contidas na vesícula acrossômica têm por função perfurar as membranas do óvulo na fecundação. (Fig. 6.9) Dentre tantas funções, o complexo golgiense também é responsável pela produ- ção da organela citoplasmática denominada lisossomo, como veremos a seguir. Figura 6.9 Representação esquemática da diferenciação do espermatozoide. À medida que a espermátide se transforma em espermatozoide, as cisternas do complexo golgiense acumulam enzimas; estas se fundem e originam a vesícula acrossômica, localizada na extremidade da cabeça do espermatozoide. (Representação sem escala, cores-fantasia.) EspErmátidE mitocôndrias Núcleo Complexo golgiense Centríolo retículo granuloso Vesícula acrossômica Crescimento do centríolo Figura 6.8 Representação esquemática da localização e da estrutura de uma célula acinosa. O pâncreas contém inúmeras bolsas, denominadas ácinos pancreáticos, cujas paredes são constituídas por células secretoras de enzimas digestivas (células acinosas). (Representação sem escala, cores-fantasia.) Diversas substâncias de natureza proteica, além das enzimas digestivas já mencionadas, passam pelo complexo golgiense para ser secretadas; é o caso de certos hormônios e de subs- tâncias mucosas, como as produzidas por nossas vias respiratórias. (Fig. 6.8) R ep ro d uç ão p ro ib id a. A rt .1 84 d o C ód ig o P en al e L ei 9 .6 10 d e 19 d e fe ve re iro d e 19 98 . 179 C a p ít u lo 6 • C it o p la sm a e o rg a n e la s ci to p la sm át ic a s 3 Lisossomos Lisossomos (do grego lise, quebra) são bolsas membranosas que contêm dezenas de tipos de enzimas digestivas, capazes de digerir grande variedade de substâncias orgânicas. Dentre as enzimas lisossômicas, destacam-se as nucleases, que digerem ácidos nucleicos (DNA e RNA), as proteases, que digerem proteínas, as fosfatases, que atuam removendo fosfatos de nucleo- tídios, de fosfolipídios e de outros compostos, além de enzimas que digerem polissacarídios e lipídios.Células animais podem conter centenas de lisossomos. A membrana lipoproteica dos lisossomos é capaz de bombear ativamente íons H1 e C,2 do citosol para o interior da organela. Esses íons formam ácido clorídrico (HC,), o que faz o interior do lisossomo ter um pH ácido, em torno de 4,8, ideal para a ação das enzimas lisossômicas. Essas enzimas praticamente não atuam em pH neutro, como o do citosol e o do líquido que banha as células. Assim, se eventualmente um lisossomo se rompe e libera suas enzimas no citoplasma, não chega a haver autodigestão da célula, pois o pH do citosol gira em torno de 7,0 e 7,3. Um mistério ainda não desvendado pelos cientistas é como a membrana do lisossomo consegue se proteger de ser digerida por suas próprias enzimas. Os lisossomos recém-produzidos pelo complexo golgiense vagam pelo citoplasma até se fundir a bolsas membranosas contendo materiais a serem digeridos. Enquanto essa fusão não ocorre, eles são denominados lisossomos primários, pois ainda não iniciaram sua atividade de digestão. Quando se fundem a bolsas membranosas com os materiais que serão digeridos e suas enzimas entram em ação, eles passam a ser chamados de lisossomos secundários. Lisossomos podem digerir material capturado do exterior por fagocitose ou por pinocitose, o que constitui sua função heterofágica, ou digerir partes desgastadas da própria célula, o que se denomina função autofágica. (Fig. 6.10) Figura 6.10 Acima, representação esquemática das funções heterofágica e autofágica dos lisossomos. (Representações sem escala, cores-fantasia.) (Baseado em Campbell, N. e cols., 1999.) À esquerda, micrografia de um corte de célula animal ao microscópio eletrônico de transmissão em que se veem lisossomos secundários (Li), alguns deles digerindo partes da própria célula (aumento 8.0003). Mi Mi Li LiLi REG 1um lisossomo secundáriolisossomo primário Material sendo fagocitado Fagossomo Vacúolo autofágico Mitocôndria inativa englobada bolsas membranosas lisossomos primários Pinossomo Material englobado por pinocitose
Mais conteúdos dessa disciplina
 17458799530346507944509115576623
17458799530346507944509115576623- 6ef1c059884e4a972df11775fb3977e994656cb9073051dda4a3894d3eb8b77de775c5486176acf38e40900cd5b01a52725ddf5c7278b61289e1b8773d3a8b0b
- Unidade 3 - Núcleo celular, ciclo e divisão celular, gametogênese - Organização cromossômica e o estud
- Screenshot_2025-04-28-17-46-16-702_com.google.android.googlequicksearchbox-edit
- Genetica 4 - Resumo
- Relatório de aula prática Ciências moleculares e celulares (RESOLVIDO)
- 17458589525582368212481591049291
- 17458589128633327372724178046103
- 17458588196395809994373941301920
- relatorio biologia
- bases de biologia celular e genética
- Prova - Análises Clínicas Microbiológicas e Imunólogicas
- Marcar para revisão A membrana mitocondrial externa possui em sua composição proteínas muito importantes. Entre elas estão as proteínas que formam ...
- 6 Marcar para revisão (Urca, 2012) Os biólogos sabem que na evolução da vida antes da galinha veio o ovo amniótico e antes da origem do DNA houve o...
- 5 Marcar para revisão Os filamentos intermediários são formados por subunidades alongadas, fibrilares e pequenas. Estes filamentos se assemelham a ...
- Qual alternativa correta o que sao celulas
- Em relacáo ao processo de diferenciacáo celular.
Assinale a afirmativa correta:
O a mudanga na expressáo génica ndo interfere na especializacáo cel...
- O retículo endoplasmático rugoso (granular) apresenta-se muito desenvolvido em células neurais, principalmente em neurónios piramidais do hipocampo...
- A organela celular responsável pela producáo de energia, através da oxidacáo da glicose é:
A organela celular responsável pela producáo de energia,...
- A maioria das organelas citoplasmáticas das células eucariontes possuem uma bicamada lipídica semelhante á bicamada da membrana plasmática. Parte d...
- A membrana plasmatica apresenta, em sua superfície, moléculas especiais que permitem a célula detectar outras substáncias presentes no meio externo...
- Costuma-se dizer que as células sáo formadas por membrana, citoplasma e núcleo. Entretanto, náo sáo todas as células que apresentam um núcleo defin...
- A Respiração Celular é um importante processo para produção de ATP, mas existe uma outra maneira de produzi-lo, a beta-oxidação.
Sobre a beta-oxida...
- O retículo endoplasmático é uma organela exclusiva de células eucariontes, que é formada por interconexões de vesículas achatadas e túbulos membran...
- Considere as afirmativas a seguir sobre as propriedades do código genético.
Assinale a alternativa correta.
I. Existem vinte e quatro tipos de amin...
- Alterações Genéticas Humanas
- resumao genetica provas online estacio
Conteúdos escolhidos para você





Perguntas dessa disciplina
Mais conteúdos dessa disciplina
- 17458799530346507944509115576623
- 6ef1c059884e4a972df11775fb3977e994656cb9073051dda4a3894d3eb8b77de775c5486176acf38e40900cd5b01a52725ddf5c7278b61289e1b8773d3a8b0b
- Unidade 3 - Núcleo celular, ciclo e divisão celular, gametogênese - Organização cromossômica e o estud
- Screenshot_2025-04-28-17-46-16-702_com.google.android.googlequicksearchbox-edit
- Genetica 4 - Resumo
- Relatório de aula prática Ciências moleculares e celulares (RESOLVIDO)
- 17458589525582368212481591049291
- 17458589128633327372724178046103
- 17458588196395809994373941301920
- relatorio biologia
- bases de biologia celular e genética
- Prova - Análises Clínicas Microbiológicas e Imunólogicas
- Marcar para revisão A membrana mitocondrial externa possui em sua composição proteínas muito importantes. Entre elas estão as proteínas que formam ...
- 6 Marcar para revisão (Urca, 2012) Os biólogos sabem que na evolução da vida antes da galinha veio o ovo amniótico e antes da origem do DNA houve o...
- 5 Marcar para revisão Os filamentos intermediários são formados por subunidades alongadas, fibrilares e pequenas. Estes filamentos se assemelham a ...
- Qual alternativa correta o que sao celulas
- Em relacáo ao processo de diferenciacáo celular.
Assinale a afirmativa correta:
O a mudanga na expressáo génica ndo interfere na especializacáo cel...
- O retículo endoplasmático rugoso (granular) apresenta-se muito desenvolvido em células neurais, principalmente em neurónios piramidais do hipocampo...
- A organela celular responsável pela producáo de energia, através da oxidacáo da glicose é:
A organela celular responsável pela producáo de energia,...
- A maioria das organelas citoplasmáticas das células eucariontes possuem uma bicamada lipídica semelhante á bicamada da membrana plasmática. Parte d...
- A membrana plasmatica apresenta, em sua superfície, moléculas especiais que permitem a célula detectar outras substáncias presentes no meio externo...
- Costuma-se dizer que as células sáo formadas por membrana, citoplasma e núcleo. Entretanto, náo sáo todas as células que apresentam um núcleo defin...
- A Respiração Celular é um importante processo para produção de ATP, mas existe uma outra maneira de produzi-lo, a beta-oxidação.
Sobre a beta-oxida...
- O retículo endoplasmático é uma organela exclusiva de células eucariontes, que é formada por interconexões de vesículas achatadas e túbulos membran...
- Considere as afirmativas a seguir sobre as propriedades do código genético.
Assinale a alternativa correta.
I. Existem vinte e quatro tipos de amin...
- Alterações Genéticas Humanas
- resumao genetica provas online estacio